DICCIONARIO MÉDICO
Apoptosis
La apoptosis es un proceso de muerte celular programada por el cual las células del organismo se autodestruyen de forma ordenada, sin desencadenar inflamación en el tejido circundante. Interviene en el desarrollo embrionario, la regulación del sistema inmunitario y el mantenimiento del equilibrio celular en los tejidos adultos. Se estima que un adulto pierde entre cincuenta y setenta mil millones de células al día por esta vía. Designa la vía de muerte celular en la que la propia célula activa un programa interno de desmantelamiento. En la necrosis, la agresión viene de fuera y la membrana celular se rompe, vertiendo el contenido al medio y provocando inflamación. Nada de eso ocurre aquí. La célula apoptótica se encoge, su cromatina se condensa, el núcleo se fragmenta y los restos quedan envueltos en pequeñas vesículas rodeadas de membrana. Esas vesículas, los llamados cuerpos apoptóticos, son reconocidas con rapidez por macrófagos y otras células con capacidad fagocítica, que las eliminan antes de que lleguen a liberar señales proinflamatorias. Procede del griego antiguo ἀπόπτωσις (apóptōsis), formada por ἀπό (apó, «desde» o «lejos de») y πτῶσις (ptōsis, «caída»). Hipócrates la empleaba para referirse a la caída de fragmentos óseos en las fracturas, y Galeno la aplicó a la caída de las costras de una herida. Ninguno de los dos la asoció con la muerte celular, un concepto que no existía en la Antigüedad. El término reapareció en la literatura médica en 1972, cuando los patólogos John F. R. Kerr, Andrew H. Wyllie y Alastair R. Currie publicaron en el British Journal of Cancer el artículo que definió por primera vez esta forma de muerte celular como un fenómeno biológico con entidad propia, distinto de la necrosis. Fue James Cormack, catedrático de griego en la Universidad de Aberdeen, quien les sugirió rescatar la palabra: la imagen de las hojas que se desprenden de un árbol en otoño le parecía un paralelo exacto de lo que ocurría bajo el microscopio. Kerr había empezado a observar el fenómeno años antes, en Brisbane, estudiando la muerte de hepatocitos tras isquemia. La célula puede recibir la orden de morir desde dentro o desde fuera. En la vía intrínseca (también llamada mitocondrial), el daño al ADN, el estrés oxidativo o la privación de factores de supervivencia alteran el equilibrio entre proteínas de la familia Bcl-2. Cuando predominan las proteínas proapoptóticas, la membrana externa de la mitocondria se permeabiliza y libera al citoplasma una molécula clave: el citocromo c. Su presencia en el citoplasma activa una cascada de enzimas proteolíticas denominadas caspasas, que desmontan la célula desde dentro cortando proteínas estructurales, de reparación y de señalización. En la vía extrínseca, la señal se inicia en la superficie celular. Ciertos receptores de la membrana (los receptores de muerte, como Fas) reciben una señal de un linfocito u otra célula del sistema inmunitario, y transmiten esa instrucción al interior mediante la activación directa de caspasas iniciadoras. Las dos vías no funcionan como compartimentos estancos; hay puntos de cruce entre ambas, de modo que una señal extrínseca puede amplificarse reclutando la maquinaria mitocondrial. La convergencia final, en cualquier caso, ocurre siempre en las mismas caspasas ejecutoras (sobre todo las caspasas 3 y 7), que son las responsables de los cambios morfológicos visibles al microscopio. Durante la formación del embrión, la apoptosis esculpe estructuras. Un ejemplo clásico es la separación de los dedos: las células del tejido interdigital se eliminan por apoptosis para que los dedos queden individualizados. En el sistema nervioso embrionario, el proceso es todavía más llamativo, porque se producen neuronas en exceso y las que no consiguen establecer conexiones sinápticas funcionales mueren. El resultado neto es que el organismo se construye eliminando tanto como creando. Sydney Brenner, H. Robert Horvitz y John E. Sulston recibieron el Premio Nobel de Fisiología o Medicina en 2002 por haber descifrado la genética de este proceso en el nematodo Caenorhabditis elegans, un gusano de apenas un milímetro en el que se eliminan exactamente 131 de las 1.090 células somáticas generadas durante el desarrollo. Cada una de esas 131 muertes está genéticamente determinada, lo cual permitió identificar los genes ced-3, ced-4 y ced-9, cuyos equivalentes humanos (las caspasas, Apaf-1 y Bcl-2, respectivamente) gobiernan también la apoptosis en nuestros tejidos. En el adulto, la apoptosis mantiene el equilibrio entre proliferación y eliminación celular. Las células del epitelio intestinal, por ejemplo, se renuevan cada pocos días: las nuevas ascienden desde las criptas y las viejas, al llegar a la punta de la vellosidad, entran en apoptosis y se desprenden hacia la luz del intestino. El sistema inmunitario depende igualmente de este mecanismo. Alrededor del 95 % de los linfocitos T que se generan en la médula ósea mueren por apoptosis en el timo antes de ser liberados a la circulación, porque han mostrado tendencia a reaccionar contra tejidos propios. Sin esa purga, el riesgo de autoinmunidad sería considerable. Cuando el equilibrio se inclina hacia uno u otro lado, aparecen consecuencias. Una apoptosis insuficiente permite que células con ADN dañado continúen dividiéndose, lo que favorece la carcinogénesis. De hecho, la evasión de la apoptosis es uno de los rasgos que comparten las células de la mayoría de las neoplasias malignas: mutaciones en el gen supresor p53 o sobreexpresión de proteínas antiapoptóticas como Bcl-2 anulan la señal de muerte y permiten que el tumor crezca. En el extremo contrario, un exceso de apoptosis destruye células que el organismo necesita. Se ha documentado una pérdida neuronal excesiva mediada por apoptosis en procesos neurodegenerativos, y la eliminación prematura de linfocitos contribuye a la inmunodeficiencia que acompaña a ciertas infecciones. Que la misma vía de muerte celular pueda estar alterada tanto por defecto como por exceso ilustra hasta qué punto su regulación es un acto de precisión continua. Fue, precisamente, la distinción entre apoptosis y necrosis el eje del artículo de Kerr, Wyllie y Currie en 1972. La necrosis es pasiva: la célula se hincha, pierde la integridad de su membrana y vierte su contenido al espacio extracelular, lo que atrae células inflamatorias y daña a las vecinas. La apoptosis es activa y limpia. Los restos se empaquetan en cuerpos apoptóticos que los fagocitos retiran sin generar inflamación. Con los años se han descrito otras modalidades de muerte celular regulada. La necroptosis comparte con la necrosis el aspecto morfológico (hinchazón, rotura de membrana), pero está controlada por vías de señalización definidas, no es accidental. La piroptosis, mediada por caspasas inflamatorias, genera poros en la membrana y libera citocinas proinflamatorias: su función es dar la alarma ante infecciones intracelulares. La autofagia, en sentido estricto, no siempre es una forma de muerte; es un mecanismo de reciclaje por el que la célula degrada componentes propios dentro de lisosomas, y solo en ciertas circunstancias conduce a la muerte celular. Estas categorías no son del todo estancas, porque comparten componentes moleculares y una misma célula puede transitar de una vía a otra según las señales que reciba. Del griego antiguo ἀπόπτωσις (apóptōsis): ἀπό, «desde», y πτῶσις, «caída». Hipócrates la usó para la caída de fragmentos óseos; Galeno, para la caída de las costras. En 1972, el catedrático de griego James Cormack, de la Universidad de Aberdeen, sugirió el término a los patólogos Kerr, Wyllie y Currie porque la imagen de las hojas desprendiéndose de un árbol les pareció la metáfora perfecta del fenómeno que observaban al microscopio. Un dato curioso: la palabra ya había aparecido en un diccionario médico español de 1878, escrito por J. Cuesta y Ckerner, pero con un significado completamente distinto: «acto de aflojar un vendaje». Durante décadas se usaron como sinónimos, pero hoy se reconoce que la muerte celular programada abarca más formas que la apoptosis. La necroptosis y la piroptosis, por ejemplo, también están reguladas genéticamente y no son accidentales. La apoptosis es la modalidad más estudiada y la que presenta el conjunto de cambios morfológicos más característico (condensación de cromatina, fragmentación nuclear, cuerpos apoptóticos), pero no es la única. En un adulto sano, entre cincuenta y setenta mil millones. Eso equivale, aproximadamente, al peso de todo el cuerpo renovado a lo largo de un año. En un niño de entre ocho y catorce años la cifra es menor, entre veinte y treinta mil millones diarios. Una célula que acumula daño en su ADN debería entrar en apoptosis antes de dividirse. Cuando ese mecanismo falla (por mutación de p53, sobreexpresión de Bcl-2 u otros defectos), la célula dañada sigue dividiéndose y puede dar origen a una neoplasia. La evasión de la apoptosis es, de hecho, uno de los rasgos que la investigación oncológica reconoce como compartido por la mayoría de los tumores malignos. Si desea ampliar conceptos vinculados a la apoptosis, puede consultar las siguientes definiciones del Diccionario médico:Qué es la apoptosis
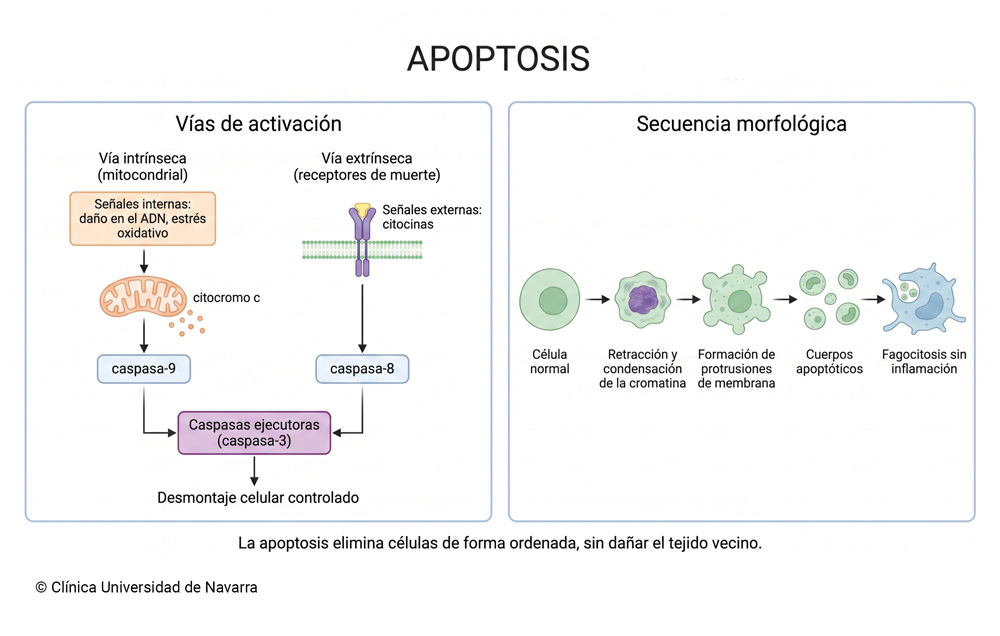
Vía intrínseca y vía extrínseca
Funciones en el desarrollo y la homeostasis
Apoptosis y enfermedad
Diferenciación con la necrosis y otras formas de muerte celular
Preguntas frecuentes
¿De dónde viene la palabra apoptosis?
¿Es lo mismo apoptosis que muerte celular programada?
¿Cuántas células mueren por apoptosis cada día?
¿Qué relación tiene la apoptosis con el cáncer?
Referencias
Entradas relacionadas en el diccionario
Infografías realizadas con https://BioRender.com
© Clínica Universidad de Navarra 2026